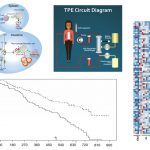
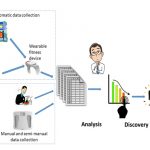
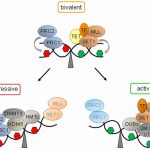
This gallery contains 6 photos.
By Vince Giuliano 9-27 -2023 Preface and a personal note In April of this year, I experienced a breakthrough in understanding human aging. This was a key event for me, coming after 15 years of full-time study and writing 600 … Continue reading